
Эубактерии
Порядок эубактерии (EUBACTERIALES)
В строгом смысле к истинным бактериям относят одноклеточных бактерий с ригидными клеточными стенками, гетеротрофов, перемещающихся (если обладают подвижностью) с помощью жгутиков. В эту группу входит преобладающее количество прокариотных микроорганизмов.
Клетки эубактерий очень малы и вместе с тем сильно различаются по размерам: от карликовых форм (диаметром около 0,125 мкм) до гигантов, длиной до 10 мкм (Ghromatium okenii). В природных субстратах (илах, почвах) можно встретить стебельковых бактерий, достигающих в длину вместе со стебельком 100 мкм и более. Следовательно, гиганты превышают карликов по размерам в 100 и 1000 раз. Большинство палочковидных бактерий по длине не превышают 5 мкм, по толщине 1 мкм. У многих псевдомонад диаметр клетки равен 0,4—0,7 мкм, у ряда почвенных форм — 0,2—0,3 мкм. 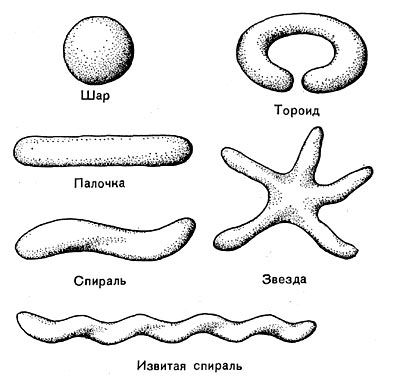
Разнообразие форм бактериальных клеток невелико. Основные формы бактерий — это шары (кокки), палочки (прямые, изогнутые или извитые), тороиды и звезды (рис. 14). Клетки многих видов неспорообразующих и спорооб-разующих бактерий выглядят как палочки. Спириллы имеют форму спирали, и ряд близких видов — форму тороида.
Поверхность клеток бактерий имеет часто придаточные структуры. Наиболее распространенными являются жгутики и ворсинки (фимбрии, нитевидные придатки клеток).
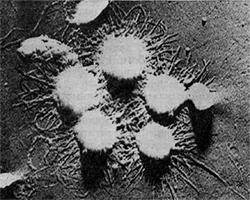
Фимбрии (или ворсинки) короче и тоньше жгутиков, имеют вид жестких прямых нитей и, покрывая всю клетку, создают ворсистую поверхность (рис. 15, 16). Длина фимбрий от 0,3—4 мкм, ширина — 0,01 мкм. Иногда они сливаются в комки, придавая неопрятный вид клетке; в других случаях поверхность клеток покрыта войлокообразным чехлом, состоящим из сплетений тонких нитей. Наряду с фимбриями можно видеть капсульный материал, как бы застрявший между нитями фимбрий, выполняющих роль арматуры. Нередко можно встретить лентовидные придатки у клеток. В среде обитания фимбрий выполняют прикрепительные функции, обеспечивая прикрепление клеток к частицам почвы, органических остатков, объединение клеток в группы (рис. 16). Есть сведения о том, что по коротким нитям, соединяющим две клетки и имеющим внутри канал, происходит обмен наследственной информацией. Среди побочных функций фимбрий — защита клетки от нападения паразитов, так как успех атаки паразитов обеспечивается непосредственным контактом с поверхностью клетки хозяина.
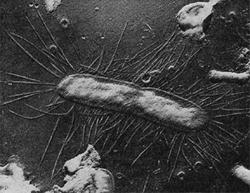
Изучение химической природы ворсинок встретилось со значительными трудностями, так как потребовалось их отделить от клетки, «побрить» клетку. К счастью, для исследователей это оказалось возможным проделать в дезинтеграторах — приборах, разрушающих клетки. При этом ворсинки обрываются, после чего их отделяют на центрифугах и накапливают. Как оказалось, они состоят из белка, обладают гидрофобностью (водоотталкивающими свойствами) и локализованы на клеточной стенке, представляя собой как бы «взлохмаченную» стенку клеток.
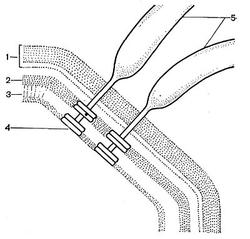
Другим типом придатков, свойственным многим бактериям, являются жгутики. По структуре они представляют собой гомогенную белковую нить или плотно свернутую косу из 2—3 нитей. Длина жгутиков 3—12 мкм, толщина — 0,02 мкм. Жгутики волнообразно изогнуты и могут быть расположены по всей поверхности клетки (перитрихиально) либо на полюсах (монотрихиально). Они составляют до 2% сухой массы бактерии. Жгутики являются органом движения бактерий, хотя известны случаи существования бездействующих (парализованных) жгутиков. Прикрепление жгутиков к клетке связано с цитоплазматической мембраной (рис. 17, 18). Основание жгутика прикреплено к специальной мембране. При делении у монотрихов дочерняя клетка приобретает полярный жгутик в том месте, где завершилось деление (рис. 19,7). В случае перитри-хиального жгутикования новые и ранее существовавшие жгутики распределяются равномерно (рис. 19,2). 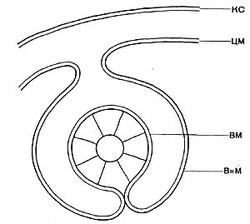
У многих микроорганизмов жесткая клеточная стенка окружена слоями слизистого материала, образующего капсулы разной плотности и толщины (рис. 20). Основными компонентами капсул являются вода и полимеры — полисахариды. Особенно обильное образование слизи наблюдается у многих микроорганизмов в тех случаях, когда среда содержит сахарозу. Молочнокислая бактерия Leuconostoc mesen-teroides за несколько часов превращает раствор в студень — декстран, который используют как заменитель крови, для повышения вязкости водных растворов. Препарат из декстрана — сефадекс — применяется в лабораториях в качестве «молекулярного сита» для разделения веществ с большой молекулярной массой.
Основным структурным компонентом клетки бактерий является клеточная стенка, которая придает протопласту микроба характерную для него форму. Когда целостность стенки нарушается, образуется сферический протопласт (при участии внутриклеточного осмотического давления, достигающего 20—50 атм и более). Клеточная стенка составляет от 10 до 50% сухой массы бактерии. Количество материала клеточной стенки увеличивается с возрастом.
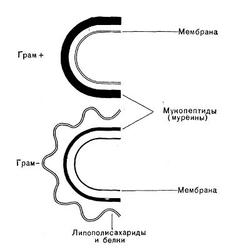
Исследования последнего десятилетия по биохимии клеточной стенки бактерий являются крупнейшими достижениями современной микробиологии, позволившими понять молекулярную организацию микробной стенки. Как уже упоминалось ранее, в зависимости от строения клеточной стенки эубактерии разделяются на 2 группы — грамположительные и грамотрицательные. В стенках грамположи-тельных бактерий очень высокое содержание гликопептидов (95% от суммы веществ стенки), однако их строение более простое, чем стенок грамотрицательных бактерий. В стенках грам-отрицательных бактерий лишь 5% гликопептидов. Этот слой покрыт рыхлым слоем белка, который, в свою очередь, прикрыт липополиса-харидным и липопротеидным слоями, пронизанными каналами (рис. 21 и 6). 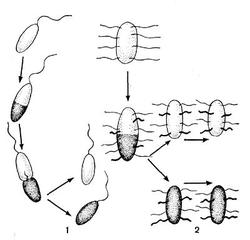
Муреиновый мешок выполняет у бактерий роль опорного каркаса. У грамположительных бактерий он многослоен, у грамотрицательных — однослоен. По существу дела, этот слой представляет собой одну молекулу, покрывающую бактериальную клетку.
Все описанные здесь особенности прокари-отных организмов являются исключительно своеобразными, свойственными представителям этого удивительного мира микроскопических существ. Внешний вид и функционально аналогичные органы (жгутики, реснички и др.) оказываются иными у эукариотных клеток.
Уникальны также структура и состав клеточных стенок бактерий. Структурные элементы стенок представляют ахиллесову пяту бактерий, так как некоторые лекарственные препараты действуют только на стенки бактерий, не затрагивая эукариотных клеток высших организмов.
Многие победы современной медицины основаны на специфичности клеточных стенок наших друзей и врагов — прокариотов.
Большие различия между эукариотами и прокариотами обнаруживаются и в мембранном аппарате клеток (в химическом составе и структуре). Так, например, у бактерий нет истинных митохондрий, играющих роль энергетических депо клеток. Их функции выполняют простые мембранные структуры (мезосомы). Особенности питания, транспорт веществ через стенку, богатейший спектр веществ, синтезируемых бактериями, особенности дифференцировки клеток, большая скорость размножения и адаптации к среде делают прокариотов истинными хозяевами и вершителями процессов круговорота веществ в биосфере.