
Подкласс растительные жгутиконосцы (фитомастигины)
ПОДКЛАСС РАСТИТЕЛЬНЫЕ ЖГУТИКОНОСЦЫ, ИЛИ ФИТОМАСТИГИНЫ (PHYTOMASTIGINA)
Многим, вероятно, приходилось видеть в природе своеобразное явление «цветения воды», когда вода в луже, пруду или озере становится изумрудно-зеленой. Причиной
«цветения» являются различные микроскопические растительные организхмы, развивающиеся в несметных количествах.
Очень часто это бывают разные представители фитомастигин.
Хламидомонады. Эти одноклеточные организмы часто являются причиной «цветения». К роду хламидомонада относится свыше 150 видов.
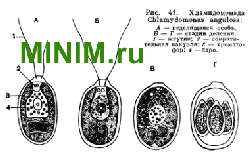
Его окраска обусловлена наличием особого пигмента, называемого гематохромом. Хламидомонады обладают резко выраженным по ложительным фототропизмом: они всегда плывут в сторону источника света. Некоторые наблюдения позволяют считать, что стигма представляет собой органоид, связанный с восприятием светового раздражения, и является по своей функции действительно клеточным глазком. В цитоплазме обычно присутствуют зерна крахмала, являющиеся продуктом фотосинтеза.
Клеточное ядро хламидомонад сферической формы и расположено примерно в середине клетки.
Размножение большинства хламидомонад происходит в неподвижном состоянии : жгутиконосец теряет жгутики, опускается на дно. Внутри оболочки происходит деление протоплазматической части клетки (протопласта) сначала на две, затем на четыре части. Через разрыв оболочки материнской клетки образовавшиеся в результате деления дочерние клетки выходят наружу, приобретают жгутики, дорастают до величины материнской клетки и выделяют оболочку.
Рассмотренный только что процесс представляет собой бесполое размножение.
Кроме бесполого размножения, у большинства хламидомонад известен половой процесс. Он сводится к тому, что вегетативные клетки становятся гаметами, которые попарно сливаются. Слияние гамет сопровождается слиянием ядер. Образовавшаяся в результате оплодотворения зигота теряет жгутики и покрывается прочной оболочкой. Внутри нее происходит деление протопласта на четыре клетки, у которых развиваются жгутики. Клетки превращаются в четыре особи хламидомонад, переходящих к свободноподвижному образу жизни.
Колониальные фитомастигины. Среди фитомастигин есть немало колониальных видов. Колонии образуются в результате бесполого размножения путем деления, при котором продукты деления остаются связанными друг с другом. Таким образом возникает как бы особь высшего порядка, включающая в себя некоторое (иногда очень значительное) число отдельных клеток, каждая из которых соответствует одной хламидомонаде. Рассмотрим некоторые наиболее широко распространенные виды колониальных фитомастигин.

Рис. 42. Колониальная фитомастпгина Gonium pectorale: 2 — жгутики; 2 — глазок (стигма); з — сократительная вакуоля; 4 — ядро. ные клетки связаны друг с другом бесцветной прозрачной студенистой массой. Эта масса образуется в результате ослизнения наружного слоя оболочек клеток.
Благодаря биению жгутиков вся колония активно плавает в воде.
Другой часто встречающейся пресноводной колониальной формой фитомастигин является эвдорина (Eudorina elegans).
Колонии этого вида, в отличие от гониумов, шаровидны и в большинстве случаев слагаются из 32 клеток (изредка число клеток колонии может быть и меньше, а именно 16 и 8). Как и у гониумов, отдельные особи колонии эвдорин связаны между собой студенистым прозрачным веществом. Диаметр колоний этого вида варьирует в довольно широких пределах (от 50 до 200 мк).
Каждая клетка колоний гониумов и эвдорин сохраняет способность к бесполому размножению, которое удается наблюдать довольно часто. К делению приступают одновременно все клетки колонии. Каждая клетка у гониума претерпевает 3, а у эвдорин — 4 последовательных, быстро следующих друг за другом деления. В результате внутри общей слизистой оболочки колонии образуются новые, дочерние колонии, число которых соответствует числу приступивших к размножению клеток. В дальнейшем слизистый чехол материнской колонии распадается, и дочерние колонии приступают к самостоятельному существованию.
Очень большой сложности достигает строение колоний у вольвокса (Volvox).
Несколько видов этого рода — частые обитатели небольших пресноводных водоемов .
Вольвоксы образуют крупные шаровидные колонии. У наиболее часто встречающегося вида Vol vox aureus диаметр колоний равен 500—850 мк, а размеры колонии Volvox globator достигают 2 мм. У Volvox aureus в состав колонии входит 500—1000 отдельных клеток (рис. 44), а у Volvox globator — до 20 тыс. Основная масса колонии состоит из студенистого вещества, образующегося в результате ослизнения клеточных оболочек. Самый наружный слой его представляет собой особый плотный кожистый слои, придающий всей колонии значительную прочность. Центральные части студенистого вещества гораздо менее плотной консистенции — они полужидкие. Отдельные клетки колонии располагаются в самом периферическом слое.
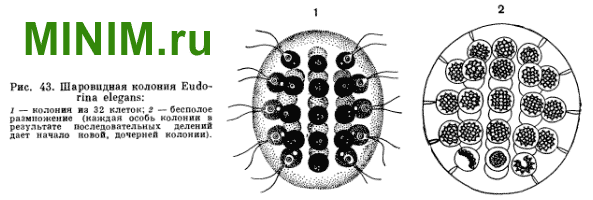
Рис. 43. Шаровидная колония Eudorina elegans: 1 — колония из 32 клеток; 2 — бесполое размножение (каждая особь колонии в результате последовательных делений дает начало новой, дочерней колонии).
Каждая клетка имеет в основных чертах такое же строение, как и одиночные хламидомонады. В колонии вольвокса отдельные особи не вполне изолированы друг от друга, они связаны между собой тончайшими цитоплазматическими мостиками.
Движение вольвокса осуществляется благодаря совместному действию жгутиков.
Несмотря на то что колония шаровидна, она обладает некоторой полярной дифференцировкой. Последняя выражается в том, что на одном полюсе стигмы (глазки) развиты сильнее, чем на другом. Движение вольвокса тоже оказывается ориентированным: он плывет вперед тем полюсом, на котором глазки сильнее развиты.
Мы видели, что у относительно просто устроенных колоний, таких, как гониум и эвдорина, все клетки способны, путем бесполого размножения давать начало новой колонии. У вольвокса дело обстоит иначе. Огромное большинство клеток колонии не способно делиться. Этим свойством обладают лишь очень немногие особи.
Число их в колонии относительно невелико: чаще всего 4—10. Они располагаются в большинстве своем в нижней части колонии (в той части ее, где глазки относительно слабо развиты). Эти клетки, называемые «вегетативными клетками размножения», удается рассмотреть лишь в относительно молодых колониях. В более старых они находятся на различных стадиях развития в дочерние колонии. Не вдаваясь в детали этого довольно сложного процесса, укажем, что дочерние колонии развиваются внутри материнской в результате многократного деления «вегетативных клеток размножения» .

Рис. 45. Небольшой участок колонии Volvox aureus (схематизированно): 1 — вегетативная особь колонии; 2 — цитоплазматический мостик; 3 — более крупная особь, в результате развития которой получаются новые, дочерние колонии.
Когда размеры дочерних колоний увеличатся настолько, что они уже не смогут поместиться внутри старой, последняя лопается и погибает, а дочерние колонии выходят наружу. Летом, в период энергичного размножения, нередко можно наблюдать, как внутри дочерних колоний, находя
Кроме бесполого размножения, у колониальных фитомастигин наблюдается и половой процесс, который протекает у различных видов по-разному. Укажем лишь, что при половом процессе в колонии образуются гаметы, в результате слияния (оплодотворения) которых получается зигота. Из зиготы путем ряда последовательных делений развивается новое поколение колоний.
Рис. 46. Разрез через колонию вольвокса с тремя дочерними колониями внутри материнской (схематизированно).