
Симметрия у животных
Расположение частей тела и органов подчинено у кишечнополостных радиальноосевой симметрии*. Симметрией называется определенный геометрический порядок в расположении сходственных частей тела.
Элементами симметрии являются точка (центр), линия (ось) и плоскость. Прекрасный пример радиально-лучевой симметрии дают радиолярии (рис.). Сходственные части тела расположены вокруг центра симметрии в радиальном направлении. Радиально-лучевая симметрия свойственна организмам, взвешенным в воде и имеющим со всех сторон одинаковую среду, в силу чего и реакция организма «одинакова во все стороны».
*Симметрия—от греческих слов sym — вместе и metron — масса, в смысле часть тела.
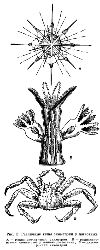
Радиально-лучевую симметрию мы находим также у колониальных фитомонадовых (вольвокс, эвдорина, пандорина и др.) и некоторых колоний многоклеточных, например у колониальной коловратки Conochilus.
Однако радиально-лучевая симметрия некоторых простейших не является самой примитивной формой строения тела. В равной мере планктонное существование также нельзя считать самой примитивной биологической формой. Самые просто организованные формы саркодовых (АтоеЫпа) имеют асимметричное строение, и, видимо, оно соответствует примитивным формам организации и поведения
(псевдоподиальная форма движения и питания). Кроме того, можно думать, что все пелагические формы существования являются вторичными производными от придонных. Асимметричное строение свойственно и инфузориям, и жгутиковым. В частности, радиолярии обладают необыкновенным богатством планов симметрии своего скелета — радиально-осевой, как гомополярной, так и гетерополярной, двусторонней, двубоковой, с обычным отклонением всех этих типов симметрии в асимметрию. Следует отметить, что в подавляющем числе случаев при этом разные формы симметрии относятся только к скелету, что же касается протоплазмы, то она, как правило, имеет асимметричное расположение включений (ядро, пульсирующие и пищеварительные вакуоли и иные включения).
Кишечнополостным, как сидячим, так и пелагическим (медузы), свойственна радиально-осевая симметрия, при которой сходственные части расположены вокруг оси вращения, причем эта симметрия может быть самого различного порядка в зависимости от того, на какой угол следует повернуть тело животного, чтобы новое положение совпало с исходным. Таким образом, может получаться 4-, 6-, 8лучевая симметрия и более, до симметрии порядка бесконечности. У радиолярий встречается радиально-осевая симметрия с одинаковыми полюсами, или, как говорят, гомополярная. У кишечнополостных — гетерополярная осевая сим26 метрия: один полюс симметрии несет рот и щупальца (оральный), другой (а б о р а л ь н ы й) служит для прикрепления (стадия полипа), или у плавающих форм несет орган чувств (ктенофоры), или ничем не вооружен (медузы).
У некоторых медуз на этой аборальной стороне образуется стебелек для прикрепления к подводным предметам (Lucernariida). Нарушение радиал ьно-осевой симметрии возникает при уменьшении числа шупалец или изменении формы ротовой щели, пищевода и разветвлений пищеварительной системы. Количество щупалец может уменьшаться до одного (Мопоbrachium), и тогда их радиальное расположение сменяется двубоковым. Глотка может сплющиваться, и тогда тоже получается двубоковая симметрия, этому способствует и образование в глотке сифоноглифов (желобок вдоль глотки).
Наибольшее усложнение радиально-осевой симметрии наблюдается у ктенофор, где, помимо 8-лучевой симметрии, в расположении отдельных частей тела и органов наблюдается 4-лучевая и двубоковая симметрия. Это весьма существенный момент, так как большинство зоологов именно от ктенофорообразных предков выводит оба ствола высших животных, как первично-, так и вторичноротых.
Гетерополярная радиальноосевая симметрия вполне соответствует образу жизни кишечнополостных — неподвижному существованию в прикрепленном положении или медленному плаванию при помощи реактивного движения.
С другой стороны, от сложного типа радиально-осевой симметрии ктенофор можно перейти к двусторонней симметрии, или, как говорят, симметрии зеркального изображения, единственного плана симметрии трехслойных животных, симметрии быстрого движения, с выработкой переднего по движению конца тела, с центральным мозговым скоплением и основными органами чувств, спинной и брюшной, правой и левой сторонами тела.
Однако живых или ископаемых свидетелей этого перехода мы не знаем. Здесь можно пользоваться только более или менее достоверными гипотезами.
В 1880 г. знаменитый эмбриолог А. Ковалевский открыл своеобразный организм — ползающую ктенофору,—названный им Coeloplana metschnikowi.
Родовым названием Ковалевский хотел показать, что этот организм объединяет в себе признаки целентерат и планарий, т. е. плоских червей. В 1886 г. другой русский зоолог — А. Коротнев, работая на о. Яве, открыл другую подобную форму, которую назвал Ctenoplana kowalewskii, также указывая в названии объединение в этом организме особенностей ктенофоры и планарий.
В настоящее время в морях Юго-Восточной Азии описан целый ряд подобных форм, объединенных в группу Platyctenidae (плоских ктенофор), но изучение их показало, что не среди них надо искать предков плоских червей, что это просто ползающие ктенофоры без предковых черт организации плоских червей.
Вопрос этот приходится решать иным путем. Возможны два варианта. По одному допущению, ктенофорообразные предки первоначально были ориентированы ротовым полюсом ко дну, а аборальным вверх. Затем они испытали расплющивание по основной оси тела и сближение орального полюса с аборальным. В дальнейшем аборальный орган чувств, зачаток мозгового скопления, должен был сместиться на тот участок сплюснутого тела, который стал передним по направлению движения. Таким образом выработались спинная и брюшная поверхности, а ротовое отверстие, как и у многих турбеллярий, осталось в средней части брюшной поверхности. Однако допущение такого пути формирования тела плоских червей должно уступить место другому. Гораздо вероятнее допустить, что ктенофорообразные предки червей ориентировались к дну боком, в таком случае у них сразу формировалась передняя по движению сторона тела, а ротовое отверстие должно было несколько сместиться по брюшной стороне вперед.
Такое допущение больше соответствует расположению нервной системы турбеллярий.

1 — движение трипаносомы при помощи ундулирующей мембраны; 2 — изгибательное движение полпхеты и 3 — рыбы. ной симметрии предков, особенно в строении нервной системы. Они сохраняют также мерцательный эпителий на поверхности тела, расположение рта на брюшной стороне и ряд других особенностей, заимствованных от целентератных предков.
Пока не сформировались рычажные конечности, основным механизмом движения оставалось изгибательное движение (рис. 4). Этот тип движения возможен при достаточно мощной мускулатуре и определенном ее расположении «пластами» вдоль всего тела. Оба эти условия сочетаются в кожно-мускульном мешке червей. При этом двигательная мускулатура составляет около половины общего объема тела, а иногда (немертины, пиявки) я значительно больше.
С образованием конечностей кожномускульный мешок распадается на отдельные мышцы. Морфологическая основа двигательной функции кожно-мускульного мешка — расположение сократительных волокон во взаимно перпендикулярном направлении. Это пласты кольцевых и продольных мышц. Еще у грегарин миофибриллы образуют систему продольных и поперечных нитей. Отростки эпителиально-мышечных клеток низших кишечнополостных также образуют слой продольных (от эктодермы) и слой кольцевых сократительных волокон. Однако у кишечнополостных количество мускулатуры невелико, кожно-мускульный мешок не формируется и движение осуществляется реактивным способом — только у ктенофор сохраняется ресничное движение, дающее, однако, при крупных размерах ктенофор очень слабый эффект, Изгибательное, волнообразное движение — весьма целесообразная форма передвижения в водной среде, однако в плотной среде грунтов, особенно морских, эта форма движения не эффективна: у животных возникает гидравлическое прямолинейное движение. При этом формируется обширная полость тела, наполненная полостной жидкостью. Количество мускулатуры в кожно-мускульном мешке уменьшается, но оно достаточно, чтобы сокращением кольцевой мускулатуры тела и перекачкой полостной жидкости вперед обеспечить продавливание хода в грунте, а затем, расширив передний конец тела и заклинив его в ходе, подтянуть заднюю часть тела сокращением продольной мускулатуры.
Принципиально тот же способ движения свойствен двустворчатым моллюскам, пробивающим ход в грунте клиновидной ногой, способной к расширению при перекачке в ее лакуны полостной жидкости, с последующим подтягиванием тела и одевающей его раковины. Интересно отметить, что отличные пловцы — головоногие моллюски — освоили реактивное движение и им не свойственно изгибательное движение тела. У их предков кожно-мускульный мешок уже распался (как и у остальных моллюсков) и была утрачена основа для создания изгибательного движения.
В водной среде возникли и членистоногие, и свойственное им движение с помощью рычажных конечностей, но, прежде чем сказать о свойственной им форме движения, следует остановиться на метамерии (сегментации) и ее происхождении.
Метамерия иногда рассматривается как своеобразный тип симметрии. При гомономной сегментации, так же как и при других типах симметрии, имеет место повторение сходственных частей тела — м ет а м е р (или сомитов) с одинаковым расположением половой системы, выделительных органов, разветвлений нервной (невросомит) и кровеносной (а нгиосомит) системы, с двумя параподиями по бокам тела, с поперечными перегородками — диссепиментами, отгораживающими сегменты друг от друга, с обособленными участками продольной и кольцевой мускулатуры (м и о с ом и т). Только в данном случае сходственные части — метамеры — расположены не вокруг точки или линии и не по обе стороны плоскости, а в линейном направлении по главной оси тела. Метамерное строение появилось вместе с развитием вторичной полости тела и кровеносной системы (рис. 5, 6). Низшие черви— сколециды — лишены настоящей метамерии (Amera), она появляется только у высших червей — аннелид — и проявляется либо в малом числе сегментов (Oligomera — мшанки, плеченогие и все вторичноротые), либо в большом (Poly mera — аннелиды и членистоногие).